Научные результаты
-
ПОДСЧЕТ СКОРОСТЕЙ МИГРАЦИИ ШИРОКОЛИСТВЕННЫХ ПОРОД
-
ХРОНО-КОРРЕЛЯЦИЯ ПАЛИНОЛОГИЧЕСКИХ ДАННЫХ ПО МИКУЛИНСКОМУ МЕЖЛЕДНИКОВЬЮ
-
РЕКОНСТРУКЦИЯ РАСТИТЕЛЬНОСТИ МИКУЛИНСКОГО МЕЖЛЕДНИКОВЬЯ
-
ЛИТЕРАТУРА
Для микулинского межледниковье в настоящее время не существует методов абсолютного датирования приемлемой точности, как например для голоцена, где они служат основой хроно-корреляции палинологических данных, которая в свою очередь необходима для восстановления динамики растительного покрова. В нашей работе хронологическая корреляция палинологических данных основана на изучении основных миграционных возможностей тех или иных видов растений. В предыдущих работах [16, 17] нами был проведен обзор и анализ представлений различных исследователей о расположении рифугиумов и основных направлениях миграционных потоков межледниковой флоры на начальных стадиях микулинского межледниковья в Европе [1, 5, 6, 8, 10, 12, 13]. В результате проведенных исследований был сделан вывод о том, что рефугиумы сыгравшие главную роль в формировании межледниковой флоры Восточно-Европейской равнины, в частности, рефугиумы неморального и кверцетального ценоэлементов [4] располагались на территории Балканского полуострова. Таким образом, миграция флоры на Восточно-Европейскую равнину при потеплении в первой половине микулинского межледниковья происходила преимущественно с юго-запада. В связи с этим, можно предположить, что наибольшее запаздывание развития лесной межледниковой флоры происходило в северных и северо-восточных регионах исследуемой территории.
ПОДСЧЕТ СКОРОСТЕЙ МИГРАЦИИ ШИРОКОЛИСТВЕННЫХ ПОРОД
Выявленные особенности миграции широколиственной флоры по территории Восточно-Европейской равнины позволяет нам предположить, что основные стадии развития растительности в северо-восточных и юго-западных регионах были не синхронны. В частности фаза максимального развития дубовых формаций в северных, восточных и северо-восточных регионах наступала существенно позже чем в юго-западных. Доказательством этому может служить сопоставление спорово-пыльцевых диаграмм разрезов, расположенных в упомянутых регионах. Так, основными общим чертами для большинства спорово-пыльцевых диаграмм Европы является обязательное присутствие на них пыльцы следующих пород – дуб, вяз, липа, граб. Максимумы пыльцы упомянутых пород последовательно сменяют друг друга в следующем порядке – дуб,вяз-липа-граб. Однако для оценки скоростей миграции широколиственных пород мы будем опираться не на чередование максимумов кривых, а на момент появления пыльцы той или иной породы в спорово-пыльцевых спектрах различных регионов. Так, в Южной Литве (разрез Лишкява [7] и др.) появление пыльцы граба на спорово-пыльцевых диаграммах происходит в начале-середине зоны липы. Однако, при продвижении к северу и северо-востоку, в разрезах Нижняя Боярщина [3], Рыбацкое [9], Рыбинск [15], Чухлома [3], Ижма [11] и др., подъем кривой пыльцы граба происходит существенно раньше и соответствует уже первой половине зоны дуба или максимуму кривой пыльцы дуба. Из этого следует, с учетом упомянутых выше направлений миграционных потоков межледниковой флоры, что скорости распространения дуба и граба с юго-запада на северо-восток не были одинаковыми. Поскольку распространение дуба на территории Восточно-Европейской равнины началось существенно раньше, чем распространение граба, очевидно, что скорость распространения граба была значительно выше.
Вследствие более высокой скорости распространения граба по территории Восточно-Европейской равнины в северных и северо-восточных регионах сокращался временной лаг между приходом упомянутых выше широколиственных пород, а наиболее восточных и северо-восточных регионов дубовые, вязовые и грабовые формации достигали одновременно (разрез Ижма [11]).
Используя этот факт можно подсчитать скорость миграции граба по территории Восточно-Европейской равнины. Это можно сделать используя исследования, проведенные Х. Мюллером по разрезу Биспинген (Германия) [22], где получены данные по длительности основных стадий развития растительности, основанные на подсчете годичных слоев озерных отложений. Согласно этим данным, фаза граба в упомянутом районе имела наибольшую продолжительность, около 4000 лет. Вместе с тем, граб был завершающим звеном в цепи смен доминантов лесных формаций, т.е. той породой, после отступания которой произошла смена неморальной растительности на бореальную. Это говорит о том, что сокращение содержания пыльцы граба в спектрах Западной Европы скорее всего означает отступание северных и восточных границ его ареала и в целом ареала широколиственной растительности. Таким образом, максимум содержания пыльцы граба в спектрах соответствует временному интервалу, когда его ареал имел наибольшую площадь распространения. Из этого также следует, что максимум содержания его пыльцы в спорово-пыльцевых спектрах был одновременным на всей территории Европы. При этом, появление граба на той или иной территории происходило не одновременно, а с увеличивающимся в восточном и северо-восточном направлениях, временным лагом (запаздыванием).
По данным Х. Мюллера [22], а также В. Загвийна [24], (последний сопоставил спорово-пыльцевую диаграмму разреза Биспинген с другими спорово-пыльцевыми диаграммами Западной Европы), от момента появления пыльцы граба в спорово-пыльцевых спектрах разрезов Западной Европы до достижения им максимального содержания прошло не более 2500 лет (рис. 1).
Чтобы использовать эти данные для подсчета скоростей миграции граба по территории Восточно-Европейской равнины необходимо учитывать, что разрез Биспинген и др. разрезы, расположенные на территории Германии, лежат вне миграционных потоков межледниковой флоры на территорию Восточно-Европейской равнины. Продвижение широколиственной флоры из своего рефугиума (Балканского п-ва) в северные регионы Восточно-Европейской равнины происходило вероятнее всего по территории Украины, Белоруссии и Прибалтики. Разрезы Южной Литвы и Германии находятся примерно на одинаковом расстоянии от Балканского рефугиума, и можно предположить, что граница ареала граба, при относительно равномерной скорости распространения, одновременно достигла регионов Германии и Литвы. Вследствие чего мы можем хронологически сопоставить диаграммы разрезов, расположенных на этих территориях. Это дает нам возможность говорить о том, что на диаграмме разреза Лишкява (Южная Прибалтика) так же как на диаграмме разреза Биспинген, от момента появления пыльцы граба до достижения ей максимума прошло около 2500 лет. За это время граница ареала граба продвинула от территории Прибалтики, до своего максимального распространения — побережья Белого моря (разрез Ижма, низовья Северной Двины) – около 1500 км. Таким образом, средняя скорость распространения граба в этом направлении, в микулинское межледниковье составляла около 600 м/год.
Vграб=S/t = 1500 000м / 2500 лет = 600 м/год.
(Vграб – скорость распространения граба по территории Восточно-Европейской равнины в микулинское межледниковье; S – расстояние от юга Прибалтики до низовьев Северной Двины (разрез Ижма); t – время, за которое граница ареала граба продвинулась с юга Прибалтики до низовьев Северной Двины (около 2500 лет)). Полученные скорости вполне сопоставимы со скоростями миграции граба, рассчитанными для голоцена (200-700 м/год) [12, 20].
При этом необходимо учесть, что скорость распространения граба не была равномерной во времени. На начальных этапах его экспансии, вероятнее всего, она была минимальной и возрастала, от начала межледниковья к его кульминации по мере установления более благоприятных условий существования данной широколиственной породы. По-видимому неравномерной была и скорость распространения граба в пространстве: естественно, что миграция граба проходила более активно в благоприятных для него условиях, т.е. в регионах, отличающихся большей океаничностью климата. Так, его продвижение вдоль восточного побережья Балтийского моря происходило существенно активнее, чем в центральные регионы Восточно-Европейской равнины.
На основании того факта, что пыльца дуба в северных регионах Восточно-Европейской равнины появляется в спектрах разрезов микулинских отложений (Ижма) одновременно с пыльцой граба, нами, также, была подсчитана и его вероятная скорость миграции для данного временного интервала. Как уже было сказано, граб достиг побережья района низовьев Северной Двины за 2500 лет. При этом, по данным Х. Мюллера [22], распространение дуба в Центральной Германии началось приблизительно на 2000 лет раньше граба. Соответственно, расстояние в 1500 км дуб преодолел за 4500 лет. Отсюда следует, что скорость распространения дуба составляла около 330 м/год.
Vдуб=S/t = 1500 000 м/4500 лет = 330 м/год.
Где Vдуб – скорость распространения дуба по территории Восточно-Европейской равнины в микулинское межледниковье; S – расстояние от разреза Лишкява (южная Прибалтика) до разреза Ижма (низовья Северной Двины); t – время, за которое граница ареала дуба продвинулась от разреза Лишкява (южная Прибалтика) до разреза Ижма (около 4500-5000 лет).
Как и в предыдущем случае, рассчитанная нами скорость миграции дуба в микулинское межледниковье вполне сопоставима с оценками скоростей его расселения в голоцене (около 300 м/год) сделанными в работах Л.Р. Серебряного [12], В. Хантли и Дж. Биркса [20].Наверх
ХРОНО-КОРРЕЛЯЦИЯ ПАЛИНОЛОГИЧЕСКИХ ДАННЫХ ПО МИКУЛИНСКОМУ МЕЖЛЕДНИКОВЬЮ
Подсчитанная скорость миграции граба позволяет более точно сопоставить спорово-пыльцевые диаграммы разрезов из различных регионов исследуемой территории. Нами были сопоставлены 4 разреза – разрез Греберн (восток Германии) [21], который был хронологически сопоставлен В. Загвийном [24] с разрезом Биспинген (центр Германии), Лишкява (Южная Литва), Рыбацкое (Ленинградская область) и Ижма (север Архангельской области) (рис. 1). Выше мы уже доказали возможность хронологического сопоставления разрезов Греберн и Лишкява. Теперь, с учетом рассчитанных выше скоростей миграции сопоставим разрезы Лишкява, Рыбацкое и Ижма.
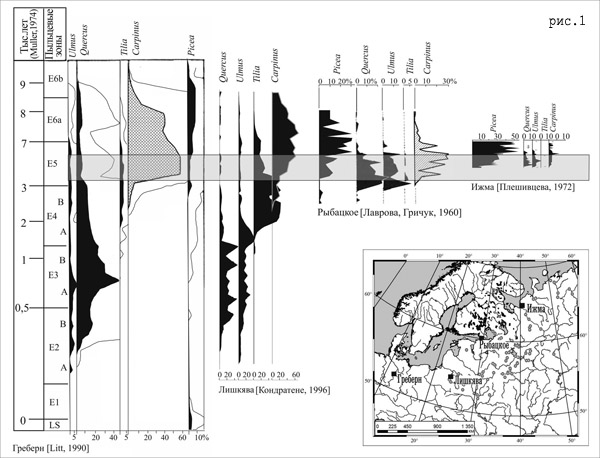
При скорости 600 м/год, граб мог расселиться из района Южной Литвы до района Ленинградской области за ~1200 лет, а до севера Архангельской области за 2500 лет. Отложив на хронологической шкале Мюллера необходимое количество лет от точки появления граба в разрезах Греберн и Лишкява, мы получим более точное время появления граба в разрезах Рыбацкое и Ижма, а следовательно – более точную хронологическую корреляцию диаграмм упомянутых разрезов.
Таким образом, при хронологическом сопоставлении спорово-пыльцевых диаграмм разрезов Греберн, Лишкява, Рыбацкое и Ижма (рис. 1) нами было выявлено, что: в северо-восточном направлении происходит сокращение продолжительности основных фаз развития широколиственной растительности и в целом сокращение времени существования широколиственных пород. Так, в Западной Европе она составляла около 7500-8000 лет, а на севере Восточно-Европейаской равнины (низовья Северной Двины) уменьшилась более чем в четыре раза и не превышала 1500-2000 лет. Сделанный вывод имеет важное значение для проведения корректной реконструкции растительного покрова на разных этапах микулинского (Эемского) межледниковья.
РЕКОНСТРУКЦИЯ РАСТИТЕЛЬНОСТИ МИКУЛИНСКОГО МЕЖЛЕДНИКОВЬЯ
Карты реконструкции растительного покрова для трех фаз развития растительности микулинского межледниковья
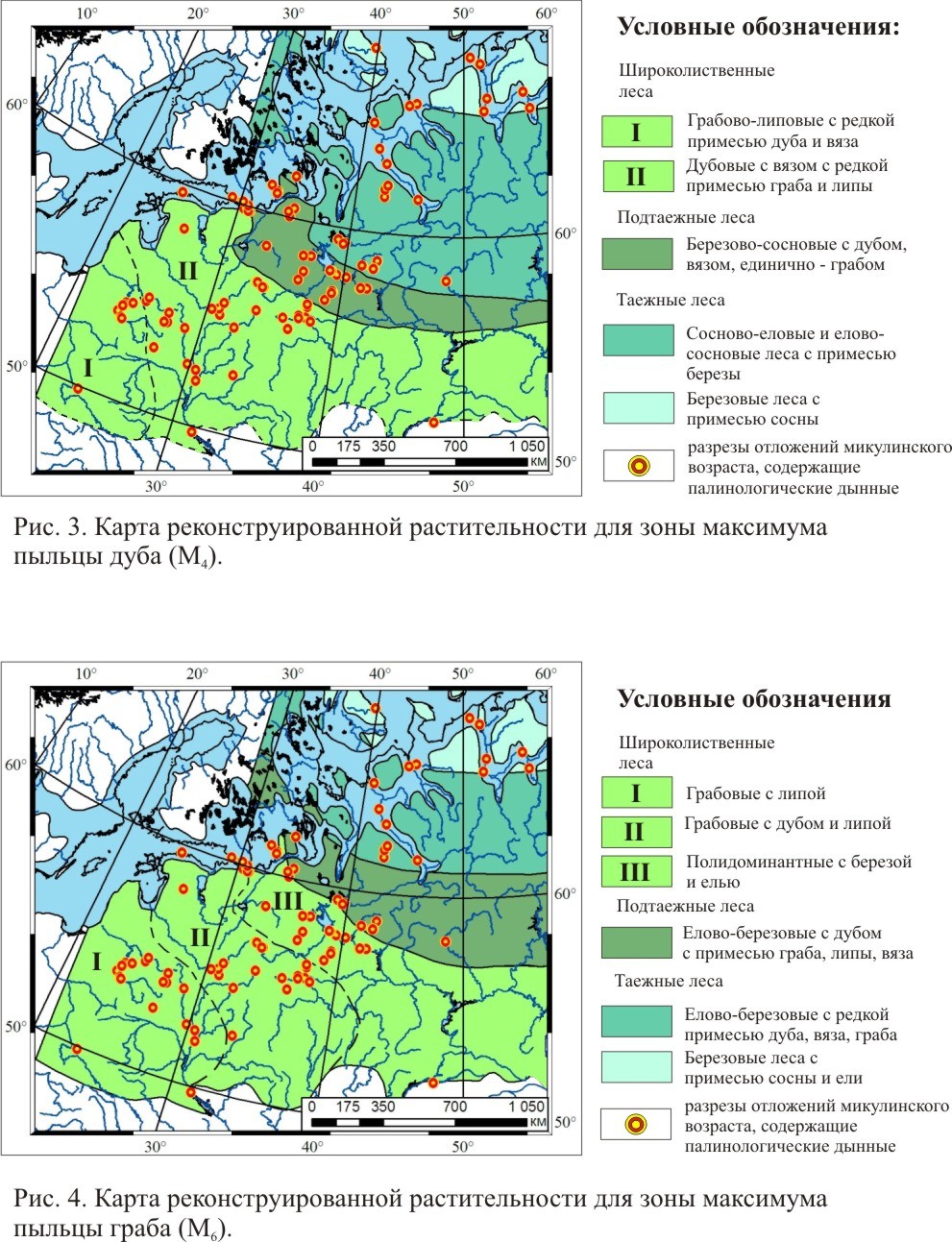
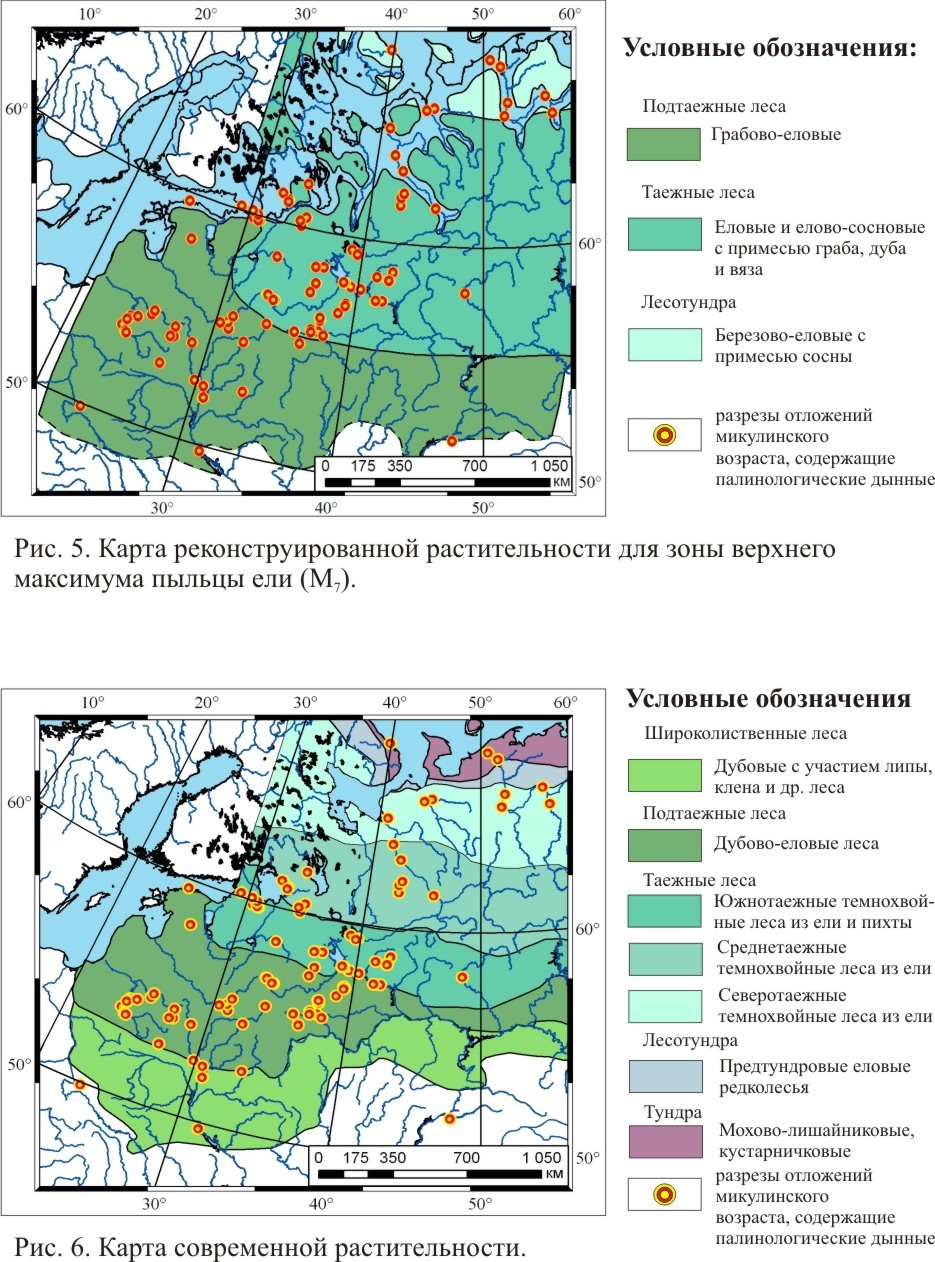
ЛИТЕРАТУРА
- Гричук В.П. К познанию процесса формирования широколиственных лесов Восточно-Европейской равнины в четвертичном периоде // Вопросы географии. 1949. Сб. 12. С. 79-96.
- Гричук В.П. Ископаемые флоры как палеонтологическая основа стратиграфии четвертичных отложений. В кн.: Рельеф и стратиграфия четвертичных отложений северо-запада Русской равнины. М.: АН СССР. 1961. С. 25-71.
- Гричук В.П. История флоры и растительности Русской равнины в плейстоцене. М.: Наука. 1989. 183 с.
- Зеликсон Э.М. Микулинская межледниковая флора Европы и ее значение для палеоклиматических реконструкций // Дисс. работа. М. 1984. 314 с.
- Казарцева Т.И. Палеогеография микулинского межледниковья Европейской части СССР (по палинологическим данным). // Автореферат диссертации на соискание ученой степени кандидата географических наук. Ленинград. 1988. 16 с.
- Клеопов Ю.Д. Основные черты развития флоры широколиственных лесов европейской части СССР. Материалы по истории флоры и растительности СССР. 1941. №1. С. 183-255.
- Кондратене О.П. Стратиграфия и палеогеография квартера Литвы, по палеоботаническим данным. // Вильнюс. Akademia-press. 1996. 214 с.
- Кожаринов А.В. Динамика растительного покрова Восточной Европы в позднеледниковье-голоцене. Автореф. к дисс. д-ра биол. наук. М.: ИЭМЭЖ РАН. 1994. 47 с.
- Лаврова М.А., Гричук М.П. Новые данные о мгинских морских межледниковых отложениях. // Доклады АНСССР. 1960. т.135. №6. с. 1472-1475.
- Морозова О.В., Кожаринов А.В. Влияние исторических факторов на распределение видового богатства флоры Восточной Европы. Известия РАН. Cер. Геогр., 2001, №5, с. 39-50.
- Плешивцева Э.С. Палинологическая характеристика опорного разреза осадков бореальной трансгрессии на северо-западе Архангельской области (район Северо-Двинской впадины) // Палинология плейстоцена. М. 1972., С. 93-104.
- Серебрянный Л.Р. Динамика распространения некоторых древесных пород на северо-западе СССР в послеледниковое время. В кн. Палинология голоцена. – М.: Наука, 1971. С.17-31.
- Удра И.Ф. Расселение растений и вопросы палео- и биогеографии. Киев, Наукова думка, 1988, 198 с.
- Федорова Р.В. Количественные закономерности распространения пыльцы древесных пород воздушным путем. // Труды ИГ АН СССР. 1952. т.52 (Материалы по геоморфологии и палеогеографии, вып.7). С. 91-103.
- Чеботарева Н.С., Макарычева И.А. Последнее оледенение Европы и его геохронология. // М: Наука. 1974. 216 с.
- Чепурная А.А. Особенности миграции некоторых широколиственных пород по территории Восточно-Европейской равнины в микулинское межледниковье // Известия РАН. Сер. Геогр. 2009. №4. С. 69-77.
- Chepurnaya A., Novenko E. “Vegetation and climate dynamics in the Central and Easten Europe (paleoenvironment reconstruction over the latitudinul transect)”; 7th European Paleobotany-Palynology Conference, Prague, 2006, p.25.
- Imbrie J., Hays J.D., Martinson D.G. et al., The orbital theory of Pleistocene climate: support from a revised chronology of the marine δ18O record // Milankovitch and Climate, Part 1. NATO ASI Series C, 126. Dordrecht: D. Reidel. 1984. pp. 269-305.
- Jessen K., Milthers V., Stratigraphical and paleontological studies of interglacial fresh-water deposits in Jutland and North-West Germany // Danm. geol. undersogelse. Raekke 2. Kobenhavn. №48. 1928.
- Huntley B. and Birks H.J.B. An Atlas of Past and Present Pollen Maps for Europe: 0-13000 BP. // Cambridge: Cambridge University Press, 1983. 667 pp.
- Litt T. Pollenanalytische Untersuchungen zur Vegetations-und Klimaentwicklung während des Jungpleistozäns in den Becken von Gröbern und Grabschüts // Altenburger Naturwissenschaftliche Forschungen. 1990. 5. P. 92-105.
- Müller H. Pollenanalytische Untersuchungen und Jahresschichtenzählung an der eem-zeitlichen Kieselgur von Bispingen-Luhe. // Geologisches Jahrbuch. 1974. A-21. P. 149-169.
- Velichko A.A., Borisova O.K., Zelikson E.M., and Nechayev V.P. Permafrost and vegetation response to global warming in North Eurasia. // In:, Biotic Feedbacks in the Global Climatic System. Will Warming Feed the Warming? (G.M. Woodwell and F.T. Mackenzie eds.). New York, Oxford: Oxford University Press. 1995. P. 134-156.
- Zagwijn W.H. An analysis of Eemian climate in Western and Central Europe // Quaternary Science Reviews. Vol. 15. 1996. P. 451-469.